In 18 years of writing Therapeutic Topics, we’ve spent a lot of time and ink considering the medicines, the mechanisms and the myriad subtleties of ocular allergic disease. In that time, therapies have evolved to a point where there are now highly efficacious treatments for most of our patients who suffer with ocular allergy. Despite this, those with the most severe, chronic allergic conjunctivitis still experience substantial disease symptomatology. These patients represent up to 30 percent of all allergic conjunctivitis sufferers, and thus constitute a significant unmet need. It seems that whether it’s the nose or the eye, our patients with chronic allergy are experiencing a simultaneous combination of immediate and delayed allergic responses, with both humoral and cellular aspects underlying their disease.
In considering how to best provide for these patients, we thought a good place to start is the beginning: the concepts of allergen priming; tolerance; and the dynamics of ocular immune responses. In this brief consideration of priming and its role in ocular allergy, we’ll ask a few key questions: What are the mechanics of priming? What role does it play in allergic conjunctivitis? What cellular mediators function in the balance between atopy and tolerance, and how can we use knowledge of these pathways to address the needs of our patients with chronic allergic disease?
Priming and Tolerance
Two events can result from antigen exposure: priming or tolerance. Priming is an event that occurs after allergen exposure; it enhances or amplifies the response of the cell or tissue to subsequent allergen exposure. This can be quantified as a lowering of threshold allergen sensitivity to achieve a response equal to that observed initially, or as a greater involvement of all cellular participants, resulting in a heightened inflammatory response.
Tolerance occurs constantly when the body’s immune system suppresses responses to cellular breakdown products. This is a necessary step to prevent constant immunogenicity from exposure to self-antigens, and autoimmune disease occurs when this homeostatic regulation breaks down. Tolerance also occurs to external antigens when a system is exposed in a particular pattern, usually to high concentrations for a short period of time, followed by constant exposure for an extended period of time. Tolerance is the physiological basis of immunotherapy.
The ebb and flow of priming and tolerance forms the foundation for the allergic reaction that is presented clinically. When exposure to allergen is repetitive, as environmental stimuli are, the reaction evolves from a discrete self-limiting response, to a series of reactions of differing onset, to a chronic inflammatory state in which the changes incurred in the tissue remain long after the first shot rings out.
Initial Response to Allergens
Allergens are proteins that have enzymatic or irritating properties that aid in penetration into the mucosa. They are internalized into dendritic cells where they are recognized by ubiquitin, a 76-residue protein highly conserved in all cellular-based organisms and present within the cytosol of DCs.1 The ubiquitin-allergen complex acts to target allergens for proteolytic fragmentation.2 These allergen fragments are transferred to the major histocompatibility complex-II on the DCs’ surface. MHC-II-antigen complexes cluster together, and are then recognized by and bound to nearby T cells.3
|
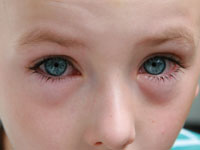
The first determinant of the priming pathway is the activation status of the DC. Activated DCs can be distinguished from resting, mature DCs by their expression of higher levels of MHCs and co-stimulatory molecules or by their production of cytokines such as IL-12 and interferon-γ. There is no longer a consideration of just mature or immature DCs, because maturation and activation are now thought to be two distinct processes. Activation of DCs involves additional stimuli or “danger signals,” changing the capacity of mature DCs from tolerogenicity to immunogenicity.5 These danger signals include IL-4, and probably other cytokines as well.
When an antigen is presented by DCs that are in a steady surveillance state, in the absence of inflammation, the result is induction of peripheral tolerance. This is a critical process for the suppression of an immune response to self. Presentation of MHC-DC clusters to T cells under steady state conditions leads initially to T-cell proliferation, followed by induction of antigen-specific non-responsiveness, called deletion or anergy.3 A second mechanism of peripheral tolerance is in the induction of antigen-specific regulatory T cells (Tregs). These mechanisms are also critical to controlling autoimmunity and are thought to be the key shutoff valves underlying the mechanisms of immunotherapy.
Another critical factor in the priming of T cells is the timing of antigen exposure. Initially short times of interaction with antigen-presenting cell clusters followed by a long antigen-independent expansion process is associated with development of atopic responses. By contrast, induction of antigen-dependent tolerance of T cells appears to require sustained contact with antigen-positive DCs beyond 72 hours. This prolonged period of exposure likely reflects an initial priming of T-cell responses by interactions with mature, activated DCs followed by a deletion of auto-reactive T cells that requires sustained interactions with DCs for longer than three days.4 With five to seven days of antigen exposure, the level of innate stimulation provided by co-stimulatory/adhesion molecules expressed by the DC is determined by the level of cytokines produced in response to various endogenous and exogenous activators (“danger” or “no danger” environment).
In the presence of adequate quantities of the “danger signal” IL-4, T cells differentiate into Th2 cells. This leads to class switching of immunoglobulins and production of IgE antibody.6 Immunoglobulin E and its high (Fcε RI) and low (Fcε RII, CD23) affinity receptors on cells are the key players in the immediate-type reactions.
Cross-talk Among Cells
Chronic allergic responses invariably include an inflammatory response, and recent evidence suggests that inflammation in peripheral tissues can lead to distant (bone marrow) induction of regulatory DCs with reduced priming ability.7 This finding highlights the importance of continual modulation of the immune pathways of priming and tolerance.
Dynamics of dendritic cell populations also play a role in allergic eye disease. In a recent series of studies, higher populations of mature dendritic cells with a reduced capacity for induction of the regulatory T-cell pathway associated with tolerance were found in vernal keratoconjunctivitis conjunctiva.8 Similar pro-inflammatory dampening of regulatory T-cell activity has also been shown in animal studies. The important and relevant discovery that the eye can locally produce the regulatory T cells critical to induction of tolerance9 highlights the plasticity of the ocular immune system, and how a local inflammatory reaction will alter not only the phenotype of bone marrow dendritic cells, as described above,7 but also immune cells in local tissues. Local mast cells are known to work in synchrony with T cells, and are usually found in close proximity to them in tissues. T cells, in turn, when activated, can degranulate mast cells, a pathway mediated by ICAM-1 binding.10 Studies of these cell-cell interactions have revealed that T cells enhance the adhesion of mast cells to vascular cells, resulting in the disruption and degradation of extracellular matrix integrity.11 It has been known for some time that the activated memory T cells that modulate mast cell function are abundant in VKC, atopic keratoconjunctivitis and giant papillary conjunctivitis.12
Eosinophils have been studied extensively in relation to priming, and are thought to be key players in the late phase and in chronic allergic conditions.13 Release of IL-5 and related cytokines prime the eosinophil, allowing for recruitment, accumulation in tissues and activation. These effects occur minutes after IL-5 exposure, and are therefore non-transcriptional processes, and the induced changes may persist for hours or days after exposure. The onset of allergy season brings changes that prime circulating eosinophils for adhesion to E-selectin and ICAM-1,14 promoting a migratory and accumulatory response in the allergic tissue. This eosinophilic priming translates clinically to more severe allergic responses.15
Eosinophils can also potentiate their own chemoattraction by providing an additional source of eotaxin-1, which, when bound to its CCR3 receptor, calls even more cells into tissues. Eotaxins have been extensively studied and are known to be elevated in chronic ocular allergic diseases.16 Recent animal studies17 have demonstrated the importance of eotaxin in allergic conjunctivitis. Eotaxin-CCR3 receptor signaling not only mediates the late phase-related eosinophilia in the conjunctiva, but is also involved in acute allergic symptoms and vascular leakage (i.e., redness and swelling). Evidence suggests this is a direct effect of eotaxin on mast cells in tissues.17 In mice deficient in eotaxin-1, the mast cell hypersensitivity response is ablated, despite normal amounts of mast cells and IgE in tissues.18 Thus, eotaxin appears to be a direct activating signal for mast cells while it also acts indirectly via binding to CCR3 receptors.
Of course, the central cell type in the acute allergic response is the mast cell. Mast cell-deficient mice have IgE and IgG antibodies that are comparable to control allergen-sensitized wild type mice, but have a minimal early phase, and no late phase or eosinophil accumulation. Both phases are fully restored by the addition of isolated naïve conjunctival mast cells, indicating that local mast cells are necessary for both the acute phase and for local, late-phase eosinophil chemoattraction (at 24 hours).19
Allergens and Priming
There is considerable research on multiple sensitivities and on how a mono-sensitized subject’s reaction can differ from a pluri-sensitized subject. This is one of the core aspects of priming. Many reports indicate that a preceding mucosal response to one allergen leads to a priming of the nasal mucosal response to another allergen. For example, in dual pet- and grass-sensitive subjects (i.e., perennial allergens), a specific nasal allergic reaction to grass in out-of-season and asymptomatic subjects was significantly more severe and more rapid than in subjects sensitive to grass alone.20
In another case, asthmatic children allergic to both mites and cockroaches were administered inhalational challenges of each, 48 hours apart. A sixfold decrease in bronchoconstriction threshold dose occurred at the second challenge, demonstrating a cross-priming effect whereby exposure to one allergen makes an individual more sensitive to a second allergen.21
Immunologically, it is known that a preexisting Th2 response to allergen can promote Th2 responses to newly encountered allergens; this is called Th2 phenotype spread.22 In an animal model, inhalation of ryegrass pollen alone does not induce sensitivity. When ovalbumin-sensitive mice are given a double, simultaneous challenge of ovalbumin and ryegrass, sensitivity to ryegrass occurs with ensuing allergic airway inflammation and progressive accumulation of specific IgE.23 This provides the mechanism behind pluri-sensitivity and also demonstrates the facility with which Th2 spread occurs in allergic subjects.
Unfortunately, this same type of phenotypic spread does not seem to facilitate immunotherapeutic approaches to allergy in pluri-sensitive subjects. The tolerance induced by specific allergen immunotherapy remains specific to that allergen, and doesn’t affect other sensitivities with regard to reductions in conjunctival threshold sensitivity and increases in IgG.24 Thus, it seems priming can be augmented by multiple sensitivities, but tolerance does not show the same trait.
Despite this limitation, immunotherapy can still be a valuable approach for attenuating a response to specific allergens in cases where the patients’ allergy is severe. This is typically done by mucosal tolerance, a dose-dependent process requiring either high doses or multiple administrations of antigen, typically administered orally. The former mode induces tolerance by clonal anergy/deletion of effector T cells, whereas the latter, based on repeated low-dose administration, causes active suppression of effector cells through regulatory T cells.
When first developed, immunotherapy was initiated slowly, with incremental increases over weeks to months. This method has been replaced by the recent “rush” and “ultrarush” immunotherapy regimens that are now widely used and considered more effective.25 In rush protocols, three hundredfold increasing doses of allergen are administered every 30 minutes, achieving the maintenance dose after three days. Ultrarush regimens give a cumulative dose of 111.1 mg in six injections, one every 30 minutes, reaching the maintenance dose in 210 minutes.
Significantly higher doses of allergen are used in immunotherapy compared to natural exposure. It’s been shown that deviation to Th2 depends on the allergen dose used to prime the corresponding experimental systems: Low allergen doses favor and high allergen doses suppress IgE production. Clinical efficacy of immunotherapy shows a strong correlation to allergen dose, with higher doses resulting in better protection. This is in direct opposition to what was thought in the early 1980s, when daily large doses of allergen were thought to lead to priming and smaller, less frequent doses were thought to lead to tolerance. This biphasic response was shown on the cellular level with basophil sensitivity to bee venom, in which low doses of allergen induced an activated state, while high doses elicited a IgE receptor desensitization, i.e., the greater the allergen/IgE cross-linking, the greater the desensitization.26
To answer the question “What is priming?” we need to look at the allergic reaction as a sum of all its parts, a complex of subsystems that respond and adapt on the fly. It’s clear that every allergic reaction in the eye, whether induced by natural or artificial challenge, not only evokes an allergic inflammatory response, but also changes the response to future exposures. The communication between mast cells, dendritic cells and T cells is intimate and constant, and thus our goal in developing treatments can be thought of as a balancing act: keeping the protective functions of the immune system intact while responding to the acute and chronic perturbations of ocular allergy. REVIEW
Dr. Abelson is a clinical professor of ophthalmology at Harvard Medical School and senior clinical scientist at the Schepens Eye Research Institute. Ms. Smith is a medical writer at Ora Inc.
1. Nandi D, Tahiliani P, et al. The ubiquitin-proteosome system. J Biosci 2006;31:137-155.
2. Wang, J, Maldonado M. The ubiquitin-proteosome system and its role in inflammatory and autommune diseases. Cell Mol Immunol 2006;3:4:255-261.
3. Kumar S, Verma A, et al. Molecular mechanisms of IgE mediated food allergy. Int Immunopharmacol 2012;13:4:432-439.
4. Dustin ML. Visualization of Cell-Cell Interaction Contacts: Synapses and Kinapses. Self Nonself 2011;2:2:85-97.
5. Tan JK, O’Neill HC. Maturation requirements for dendritic cells in T cell stimulation leading to tolerance versus immunity. J Leukoc Biol 2005;78:2:319-324.
6. Lipscomb, M, Wilder J, et al. Dendritic cells and their role in linking innate and adaptive immune responses. In: Gessani S, Belardelli F, eds. The Biology of Dendritic Cells and HIV Infection, Part 1. New York: Springer, 2007:45-84.
7. Scott NM, Ng RL, et al. Toward homeostasis: Regulatory dendritic cells from the bone marrow of mice with inflammation of the airways and peritoneal cavity. Am J Pathol 2012;181:2:535.
8. Manzouri B, Ohbayashi M, et al. Characterization of dendritic cell phenotype in allergic conjunctiva: Increased expression of Fc(epsilon)RI, the high-affinity receptor for immunoglobulin E. Eye (Lond) 2009;23:11:2099-2104.
9. Zhou R, Horai R, et al. The living eye disarms uncommitted autoreactive T cells by converting them to Foxp3(+) regulatory cells following local antigen recognition. J Immunol 2012;188:4:1742-1750.
10. Vaday G, Baram GD, et al. Cytokine production and matrix metalloproteinase (MMP)-9 release by human mast cells following cell-to-cell contact with activated T cells. Isr Med Assoc J 2000;2 Suppl:26.
11. Brill A, Baram D, et al. Induction of mast cell interactions with blood vessel wall components by direct contact with intact T cells or T cell membranes in vitro. Clin Exp Allergy 2004;34:11:1725.
12. Metz D, Bacon AS, et al. Phenotypic characterization of T cells infiltrating the conjunctiva in chronic allergic eye disease. J Allergy Clin Immunol 1996;98:3:686-696.
13. Godthelp T, Holm AF, et al. Dynamics of nasal eosinophils in response to a nonnatural allergen challenge in patients with allergic rhinitis and control subjects: A biopsy and brush study. J Allergy Clin Immunol 1996;97:3:800-811.
14. Hakansson L, Heinrich C, et al. Priming of eosinophil adhesion in patients with birch pollen allergy during pollen season: Effect of immunotherapy. J Allergy Clin Immunol 1997;99:4:551-562.
15. Yuki A, Terada T, et al. Evaluating the effects of testing period on pollinosis symptoms using an allergen challenge chamber. Allergol Int 2011;60:4:533-539.
16. Leonardi A, Jose P, et al. Tear and mucus eotaxin-1 and eotaxin-2 in allergic keratoconjunctivitis. Ophthalmology 2003;110:3:487-492.
17. Fukuda K, Kuo C, et al. The murine CCR3 receptor regulates both eosinophilia and hyperresponsiveness in IgE-mediated allergic conjunctivitis. Br J Ophthalmol 2012;96:8:1132-1136.
18. Miyazaki D, Nakamura T, et al. Ablation of type I hypersensitivity in experimental allergic conjunctivitis by eotaxin-1/CCR3 blockade. Int Immunol 2009;21:2:187-201
19. Miyazaki D, Tominaga T, et al. Conjunctival mast cell as a mediator of eosinophilic response in ocular allergy. Mol Vis 2008;14:1525-1532.
20. Toth J, Schultze-Werninghaus C, et al. Environmental priming influences allergen-specific nasal reactivity. Allergy 1998;53:12:1172-1177.
21. Koh Y, Choi J, et al. A preceding airway reaction to one allergen may lead to priming of the airway responses to another allergen. Allergy 1997;52:3:284-292.
22. Hayashi T, Gong X, et al. Induction and inhibition of the Th2 phenotype spread: implications for childhood asthma. J Immunol 2005;174:9:5864-5873.
23. Cadot P, Meyts I, et al. Sensitization to inhaled ryegrass pollen by collateral priming in a murine model of allergic respiratory disease. Int Arch Allergy Immunol 2010;152:3:233-242.
24. Dreborg S, Lee T, et al. Immunotherapy is allergen-specific: A double-blind trial of mite or timothy extract in mite and grass dual-allergic patients. Int Arch Allergy Immunol 2012;158:1:63.
25. Harvey S, Laurie S, et al. Safety of rush immunotherapy to multiple aeroallergens in an adult population. Ann Allergy Asthma Immunol 2004;92:4:414-419.
26. Chirumbolo S, Zanoni G, et al. In vitro biphasic effect of honey bee venom on basophils from screened healthy blood donors. Allergy Asthma Immunol Res 2011;3:1:58-61.



